Les chauves-souris ont évolué avec des caractéristiques uniques telles que l’écholocation laryngée et le vol, certaines étant capables de tolérer des virus tels que les coronavirus du syndrome respiratoire aigu sévère (SRAS-CoV), les CoV du syndrome respiratoire du Moyen-Orient (MERS-CoV), ainsi que les virus Marburg et Nipah. . Le développement de modèles de chauves-souris robustes à base de cellules pourrait fournir une meilleure compréhension de la manipulation et de la biologie virales des chauves-souris.
Dans une étude récente publiée sur le serveur de préimpression bioRxiv*, les chercheurs ont généré des cellules souches pluripotentes induites (iPSC) à partir de Rhinolophus ferrumequinum chauves-souris utilisant le protocole Yamanaka modifié pour établir les chauves-souris comme un roman in vivo espèce d’étude modèle.
Étude: Les cellules souches pluripotentes de chauve-souris révèlent un enchevêtrement unique entre l’hôte et les virus. Crédit d’image : Jezper / Shutterstock.com
Sommaire
À propos de l’étude
Dans la présente étude, les chercheurs étudient si les chauves-souris pourraient convenir à la production virale.
L’approche de reprogrammation Yamanaka a été utilisée sur la base de facteurs de reprogrammation tels que le gène Y-box 2 de la région déterminant le sexe, le facteur de transcription octamère 4 (Oct4), cMyc et le facteur 4 de type Kruppel (Klf4).
Des cellules de fibroblastes embryonnaires de chauve-souris (BEF) ont été isolées de R. ferrumequinum, avec les quantités de facteurs de reprogrammation modifiées pour activer et bloquer plusieurs voies de signalisation. De plus, des analyses d’immunocoloration et de séquençage de l’acide ribonucléique (ARN) (ARN-seq) ont également été effectuées.
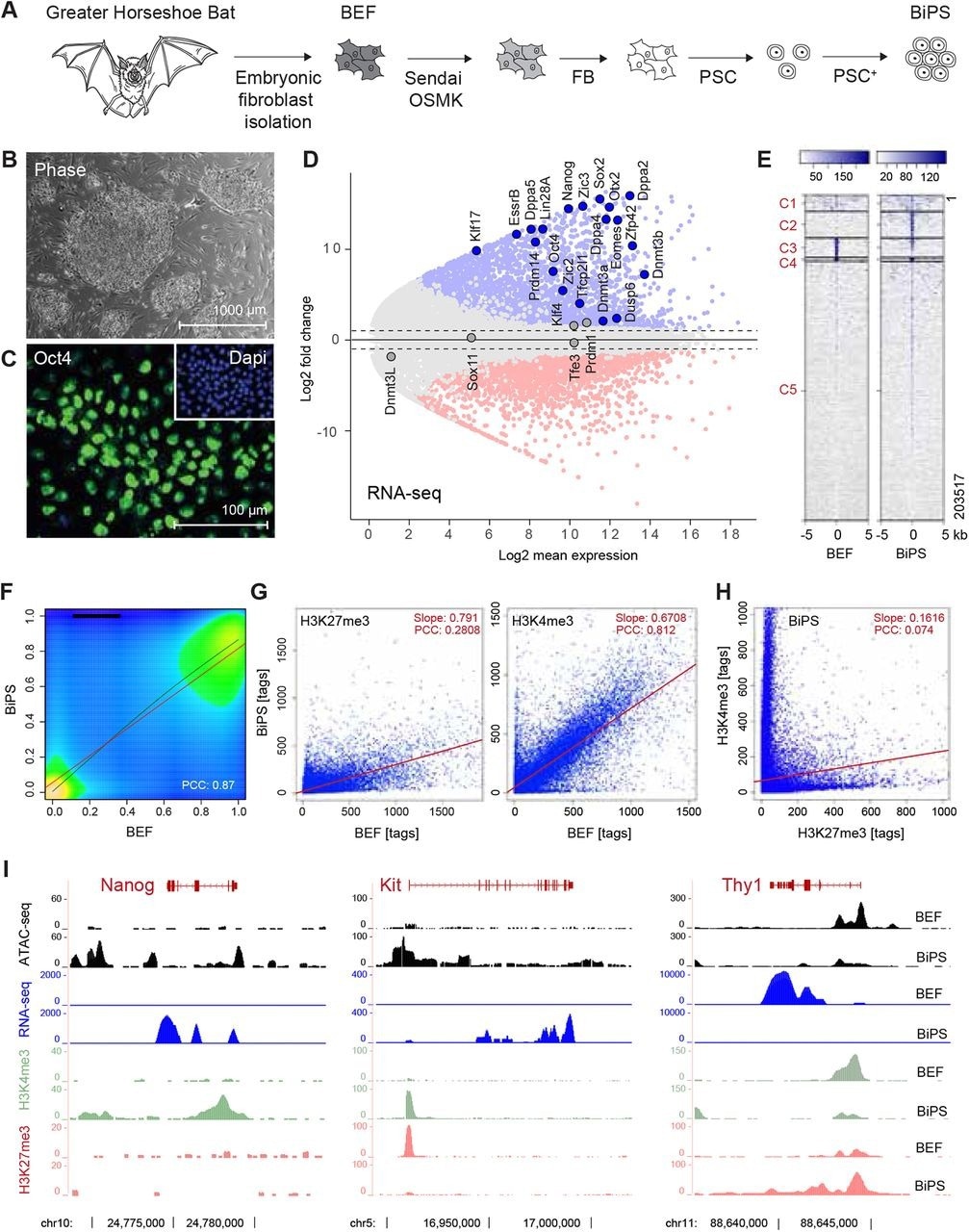
Dérivation de cellules souches pluripotentes de chauve-souris. (A) Illustration de la stratégie de dérivation de cellules souches pluripotentes de chauve-souris. BEF, fibroblastes embryonnaires ; OSMK, Oct4, Sox2, cMyc, Klf4 ; FB, milieu fibroblastique ; PSC, milieu de cellules souches pluripotentes ; PSC+, PSC avec additifs. (B) Morphologie des colonies de cellules BiPS établies cultivées sur des fibroblastes embryonnaires de souris. (C) Détection immunofluorescente d’Oct4 dans les cellules BiPS. (D) MA parcelle de données ARN-seq illustrant les différences transcriptionnelles entre le fibroblaste embryonnaire de chauve-souris (BEF) et les cellules souches pluripotentes (BiPS). Des gènes sélectionnés ayant des fonctions connues dans l’établissement ou le maintien de la pluripotence sont mis en évidence. (E) Analyse en grappes Kmean des signaux ATAC-seq obtenus à partir de cellules BEF ou BiPS. C, grappe. (F), parcelle de densité des résultats RRBS obtenus à partir de cellules BEF et BiPS. PCC, coefficient de corrélation de Pearson. (G) Nuages de points de l’état de méthylation de l’histone 3 à K4 (activation de la modification de la chromatine) ou K27 (répression de la modification de la chromatine) après ChIP-seq à partir de cellules BEF ou BiPS comme indiqué. (H) Nuage de points de H3K4me3 et H3K27me3 dans les cellules BiPS illustrant la présence de sites de chromatine bivalents dans les cellules BiPS. (I) Signaux ARN-seq, ATAC-seq et H3K4me3 ou H3K27me3 ChIP-seq de gènes sélectionnés avec des rôles connus dans la reprogrammation qui sont activés (Nanog, Kit) ou réprimés (Thy1) dans BiPS par rapport aux cellules BEF.
L’impact de la méthode de reprogrammation modifiée sur les molécules épigénétiques de chauve-souris et la chromatine a été évalué à l’aide du test de chromatine accessible à la transposase avec séquençage (ATAC-seq). Une analyse de cartographie du méthylome de l’acide désoxyribonucléique (ADN) et des analyses d’immunoprécipitation et de séquençage de la chromatine (ChIP-seq) ont également été effectuées. Les protocoles ont été optimisés pour permettre la différenciation de la chauve-souris SC dans les trois couches germinales, tandis que le test de différenciation du corps embryoïde (EB) a été effectué pour évaluer la pluripotence.
Des iPSC de chauve-souris (BiPSC) ont ensuite été injectés à des souris immunodéprimées, et des structures ressemblant à des embryons ont été créées à partir du BiPS. Le protocole d’étude a été validé en développant des cellules BiPS de l’évolution distante Chauves-souris Myotis myotis.
Le profilage comparatif des gènes transcriptionnels et l’analyse en composantes principales (ACP) ont été réalisés sur le Rhinolophus espèces de chauves-souris et espèces de mammifères phylogénétiquement différentes de souris, d’humains, de chiens, de porcs et de marmousets.
L’analyse de l’ontologie des gènes a été effectuée pour évaluer l’enrichissement génétique de pointe pour des voies biologiques spécifiques. De nouveaux pipelines ont été développés sur la base de la classification métagénomique des données séquencées en acide ribonucléique (ARN) des cellules souches (ARN-seq), de novo assemblage de contigs rétroviraux putatifs et cartographie génomique pour identifier les vraies lectures rétrovirales. De plus, des marqueurs antigéniques liés aux virus à ARN ont été explorés.
Résultats de l’étude
Un rapport de facteur de reprogrammation particulier, ainsi que le facteur de croissance des fibroblastes 2 (Fgf-2), le facteur de cellules souches (Scf), le facteur inhibiteur de la leucémie (Lif) et la forskoline ajoutés au milieu de culture ont permis une croissance BiPSC non inhibée, avec une croissance homogène et serrée colonies de chauves-souris apparaissant dans les 14 à 16 jours.
Les BiPSC ont exprimé le facteur de pluripotence Oct4, avec un taux de prolifération identique au taux de prolifération des PSC humains. La plupart des cellules contenaient 56 chromosomes et se répliquaient sans facteurs de reprogrammation exogènes ni altérations morphologiques.
Les BiPSC se sont différenciés en trois couches germinales, formant ensuite des EB et des organoïdes. L’analyse ARN-seq a montré l’expression endogène induite de gènes canoniques liés à la pluripotence comme SRY-2, Nanog et Oct4.
Cependant, le profil génétique ne correspondait pas entièrement à un état de pluripotence. Au lieu de cela, des facteurs d’état pluripotent naïfs tels que Klf4 et 17, la protéine bêta du récepteur lié aux œstrogènes (Essrb), le facteur de transcription E3 (Tfe3) et le facteur de transcription CP2 Like 1 (Tfcp2l1)]ont été exprimés. Co-exprimé Tfcp2l1/zinc finger protein (Zic2) et orthodenticle homeobox 2 (Otx2)/Tfe3 et des facteurs amorcés/naïfs ont été observés.
Des altérations de la configuration de la chromatine et de la méthylation du CpG 191 ont été observées dans le génome de la chauve-souris. Les résultats de ChiP-seq ont montré un chevauchement entre les gènes de bivalence de l’homme et de la chauve-souris, bien que certains gènes soient spécifiques à l’espèce.
Les BiPSC ont été reprogrammés transcriptionnellement et épigénétiquement. Les BIPSC étaient positifs pour les marqueurs de protéine à boîte appariée (Pax6), 213T et alpha-foetoprotéine (AFP) pour l’ectoderme, le mésoderme et l’endoderme, respectivement.
Le gène ERAS était régulé à la baisse, tandis que les gènes des hyaluronidases et des facteurs de ribosylation ADP (ARF) étaient indiscernables entre les groupes. La Rhinolophus les blastoïdes ont montré des structures embryonnaires attachées à une excroissance épithéliale trophoblastique aplatie et à une expansion de la masse cellulaire interne. Myotis les découvertes sur les chauves-souris ont indiqué que le protocole d’étude pourrait être appliqué à différentes espèces de chauves-souris.
L’analyse PCA a montré un groupe distinct de cellules souches de chauve-souris. Cependant, seuls huit gènes de pointe ont montré une sélection positive significative dans R. ferrumequinum, la plupart des gènes appartenant à des catégories inattendues. De plus, la maladie CoV était la catégorie la plus significativement enrichie dans les voies de l’encyclopédie des gènes et des génomes de Kyoto (KEGG).
Les gènes du collagène de type III alpha 1 (Col3a1) et de la mucine 1 (Muc1) ont été détectés dans les BiPSC, indiquant ainsi des adaptations génétiques spécifiques aux chauves-souris. La reprogrammation a révélé des séquences de rétrovirus endogène (ERV).
Les BiPSC contenaient plusieurs séquences endogénisées associées au virus, avec des régions homologues à l’herpèsvirus humain 4, au virus respiratoire syncytial humain et à un isolat du SRAS-CoV-2. R. ferrumequinum les séquences génomiques ressemblaient à celles du CoV 229E humain et du CoV OC43 humain.
Plusieurs sites d’intégration rétroviraux homologues à des virus tels que le virus du singe Mason-Pfizer, le virus Koala et le rétrovirus du mouton Jaagsiekte ont été identifiés. Le génome était homologue aux virus du volepox, de la variole, de la variole de l’écureuil, de la variole du singe et du syndrome des points blancs.
conclusion
Les séquences BiPSC étaient similaires aux séquences du génome viral. Ainsi, l’état de pluripotence transcriptionnellement permissif des chauves-souris pourrait être exploité pour découvrir de nouvelles séquences de virus de chauve-souris impliquées dans la physiologie des chauves-souris et leurs capacités d’hébergement de virus.
*Avis important
bioRxiv publie des rapports scientifiques préliminaires qui ne sont pas évalués par des pairs et, par conséquent, ne doivent pas être considérés comme concluants, guider la pratique clinique/les comportements liés à la santé, ou traités comme des informations établies.